鳞翅目幼虫味觉器官的形态与功能
鳞翅目幼虫味觉器官的形态与功能

鳞翅目幼虫味觉器官的形态与功能

摘要
味觉在昆虫的行为中起着关键的作用。幼虫主要依靠植物的味觉线索来探测和定位食物来源。味觉刺激可以是简单的,也可以是复杂的多分子混合物。昆虫面临的任务是破译这些味觉化合物的性质,然后必须做出适当的取食选择。通常情况下,幼虫的口器上有四种类型的两侧对称的味觉感器。侧面和中间的栓锥感器被认为是参与进食的主要器官。这些感器在取食过程中与植物汁液持续接触,可以检测到植物中存在的不同植物化学物质。味觉的感觉输入是由在这些感器中的味觉受体细胞来编码为神经脉冲模式。因此,这些味觉感受细胞形成取食选择过程的第一级,最终决定食物是被昆虫接受还是拒绝。舞毒蛾幼虫(Lymantria dispar)(鳞翅目:毒蛾科)是美国大部分地区的主要森林害虫。这些幼虫是高度多食性的取食者,取食各种树木落叶。本章讨论了与舞毒蛾幼虫有关的味觉感受的形态、取食行为和电生理。
1. 简介
味觉对动物的生存和营养至关重要。它在确定食物的适口性和提供食物腐败的早期预警信号方面至关重要。本章利用昆虫–舞毒蛾幼虫(Lymantria dispar (L.)),促进人们更好地了解味觉刺激是如何被安置在味觉器官(感器)中的受体细胞识别、编码和处理的。这些感觉器官是角质结构的,里面有味觉受体细胞。这些受体细胞构成了环境味觉信号的感觉过滤器。在昆虫中,这些感器将信息直接传递给大脑中的高级味觉处理中心,并形成取食决策过程的第一级,决定是否应该接受或拒绝食物。通常情况下,昆虫面临的任务是破译复杂的多分子混合物中的单个味觉物质,以做出适当的取食选择。为了应对不同行为或生态环境中的刺激,并区分有意义的味觉刺激,幼虫已经进化出几种不同类型的味觉感器。
植物的识别主要受位于幼虫口器上的两对感器的活动支配,即中栓锥感器和侧栓锥感器 。当幼虫进食时,这些味觉感器与植物的汁液持续接触,可以检测到植物中存在的不同化学物质(例如植物化学物质、植物次生化合物、植物化感物质)。幼虫的味觉感器为解决有关味觉系统的问题提供了一个很好的系统,因为:i)这些感器形成了一个相对简单的感觉系统,数量有限的感觉细胞介导的味觉机制;ii)这些感器很容易被实验操作,iii)这些感器内的受体细胞可以单独识别,并表现出典型的稳定的和可重复的电生理反应。
2. 化学感觉系统和感器类型
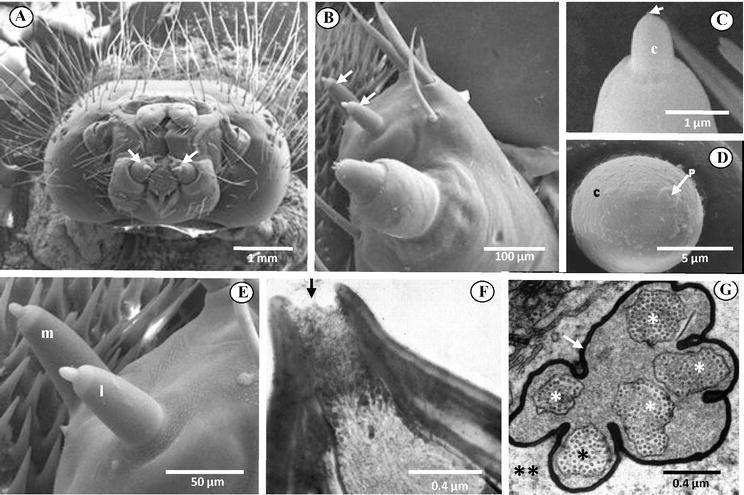
成虫拥有几种不同类型的感觉器官,监测环境中与寻找食物、产卵地、同种配偶、合适的温度和湿度水平有关的线索,并寻求保护和定位。这些感觉器官使它们能够检测到与味道、气味、触觉、声音、视觉、本体感觉以及地理、温度和湿度有关的刺激。相比之下,幼虫的感觉,如鳞翅目中的幼虫,则更为有限。例如,它们强烈地依赖味觉、触觉,可能还有短距离的嗅觉线索来选择宿主植物。鳞翅目昆虫使用各种物理和化学特征来定位植物。虽然视觉感官可以帮助幼虫到达植物附近,但这种感官的发展还不够精细,不能在识别植物方面发挥作用。昆虫的化学感觉很发达,不仅帮助单食性昆虫(只吃一种或几种密切相关的植物种类)找到它的食物,而且还帮助多食性昆虫(吃属于不同植物科的许多植物)辨别各种植物种类。化学感器位于触角和口器上(图1)。鳞翅目幼虫的头部总共有五种类型的双侧化学感器:一对触角(每个由16个神经元支配),两对位于外额叶上的中栓锥感器和侧栓锥感器(每对由8个神经元支配),一对下颚须(每个顶端表面有8个感器,每个由14-19个嗅觉和味觉神经元支配),和一对内唇(每个由三个味觉神经元支配)。
有三个主要类别的昆虫感器:(1)AP(无孔)或NP(无孔)感器,它们对机械感受或对湿度和温度感受;(2)UP(单孔)或TP(顶孔)感器含有味觉神经元,单独或与机械感受细胞一起,以及(3)MP(多孔)或WP(壁孔)感器(单壁(SW)感器和双壁(DW)感器)。通常,多孔感器是司嗅觉的,壁孔是嗅觉和/或热敏感的。中栓锥感器和侧栓锥感器单孔或顶孔感器。
3. 中栓锥感器和侧栓锥感器
昆虫的味觉被称为接触性化学感受。接触性化学感器类似于位于脊椎动物口腔中舌头上的味蕾。在鳞翅目幼虫中,味觉感器位于口器上,特别是下颚和上唇。每个下颚由一个下颚须和一个外颚叶组成。每个外颚叶上有两个细长的突起,即中栓锥感器和侧栓锥感器(图1和2),这些感器靠近幼虫口器。在取食过程中,这些感器在植物汁液进入口器或颊腔之前与之持续接触,可以探测到植物汁液中存在的不同化学物质(即植物化感物质)。在鳞翅目幼虫中,植物的识别被认为主要是由每一对的中栓锥感器和侧栓锥感器的感觉输入所介导的。因此,它们被认为是参与取食的主要感觉器官。
消除实验表明,去除烟草天蛾(Manduca sexta)的栓锥感器,导致其宿主范围扩大。消除该物种的栓锥感器,允许它在以前不能接受的植物上取食,从而扩大了这种昆虫可容忍的植物种类范围。结论是,对植物的排斥可以由中栓锥感器和侧栓锥感器介导,两种感器都参与了对不同化学物质的排斥行为。因此,这些结果支持这样的观点,即存在于每个栓锥感器中的受体细胞都参与选择性地介导阻断取食行为。明视场光镜以及透射电子显微镜显示,这些感器每个都有一个可渗透的顶端孔(单孔,UP,或终端孔,TP),通常由五个双极神经元支配,其中四个作为味觉感器,一个作为机械感器(图2)。栓锥感器之所以如此命名,是因为它看起来像一个小锥体、钉子或旋钮状结构,插入到不敏感的角质层的圆柱形突起(图1和2),因此被归类为单孔(UP)感器。
4. 单孔栓锥型感器结构
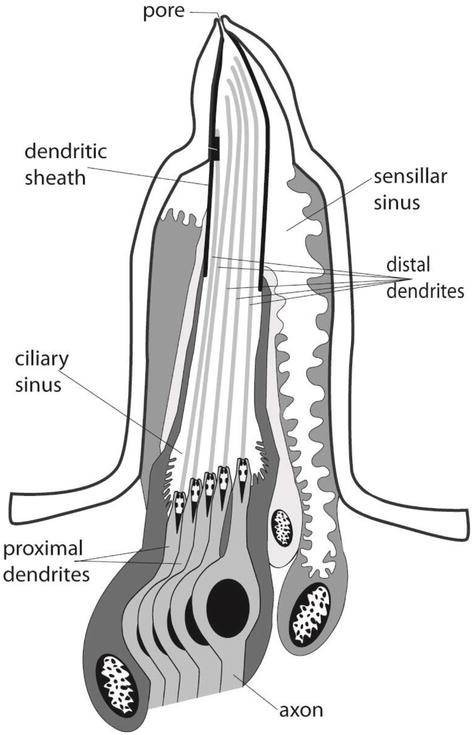
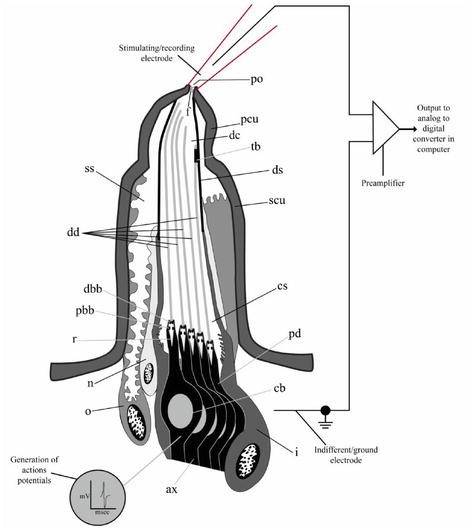
单孔栓锥感器可以采取短至中长的钉子或圆锥体的结构,插入到纤维角质层中,使其在该关节区弯曲(图1和2)。图2显示了一个栓锥感器的重建示意图。感器有一个位于顶端的单一可渗透的顶端孔,通常由五个双极神经元支配,其中四个作为味觉感器,一个作为机械感器。孔的直径约为10-200纳米,包含典型的孔管或塞子状的栅栏纤维,允许受体细胞和外部环境之间发生化学通讯。孔内纤维也可能赋予传导机制的选择性和对感器反应的特异性。四个假定的味觉神经元在感器内部的树突通道内从孔中延伸出来。一个树突鞘将树突包围起来。树突鞘从感器的顶端附近延伸到与纤毛窦的水平位置。睫状纤毛窦包裹着树突。树突鞘将树突与一个大的感觉窦完全分开。树突鞘,可能在某些区域有孔,可以使感觉窦作为离子和静息电位的储存器,正如成年果蝇的味觉所显示的那样。
第五条无分支的机械感觉树突开始于锥体基部附近,紧贴着锥体的树突鞘和角质层壁。这个树突的顶端有一个微管的堆积。这些微管在一个电子密集的基质(管状体)内相互平行,被认为是机械刺激的感觉转导的场所。树突在睫状区沿其长度的中间位置突然收缩。这一点区分了远端树突(睫状体)片段和近端树突片段。近端树突继续靠近并形成细胞体。从这一点出发,来自中栓锥感器和侧栓锥感器的轴突合并,形成外颚叶神经的侧面和内侧分支,直接投射到食道下神经节(SOG)而不发生突触连接。SOG被认为是中枢神经系统的第一级中继站。SOG还对直接参与取食过程的口器进行运动控制。各种输入(包括味觉细胞)的大部分中央处理发生在SOG,然而,由于来自中枢神经系统其他部分的输入(如额叶、嗅叶)也有助于取食行为(即宿主植物识别),目前还不清楚 “取食中心 “是否完全位于SOG。
5. 取食行为
所有的昆虫在选择食物时都有一定的选择性,以(a)一种或几种密切相关的植物物种为食(单食),(b)通常局限于某一植物科的较多宿主(寡食)或(c)取食广泛分类范围的许多植物(多食)。然而,昆虫从不以所有的植物群为食。昆虫口器上的接触性化学感器的主要功能是选择食物。当昆虫咬到植物时,一些接触性化学感器会暴露在植物的汁液中,其功能类似于脊椎动物的味觉感器,检测溶液中的化合物。然而,一些口器感器,如在鳞翅目幼虫中发现的(如在下颚和下唇须上发现的),经常在昆虫撕咬之前接触食物。当这些感器与植物表面短暂接触时,这些感器内的受体对叶子干燥表面的化合物敏感。这种拍打行为的作用是 (1)允许昆虫从感器中获得比保持接触时更持续的信息流,因为感器会变得适应;(2)允许昆虫从更大的叶子表面采样,而不是使感器保持静止。因此,通过感器拍打植物表面获得的信息会提醒昆虫避免摄入有毒的化合物,并更迅速地做出取食选择。
食物选择行为可以比作一个 “钥匙–锁 “系统,其中钥匙代表受体活性曲线。只有当这个轮廓与中枢神经系统中的模式识别区的先天标准充分对应时,才会触发特定的行为反应。当传入的感觉信息与期望的模式相差太大时,食物就会被拒绝。由SOG和其他大脑区域组成的中枢神经系统(或称之为锁)被调整为识别感觉模式。那些被识别为可接受的模式将释放取食行为,而其他模式将导致食物被拒绝。最后的决定被认为是在SOG中做出的。对于专食性昆虫来说,传入的感觉模式必须与中枢神经系统设定的某种规范更紧密地匹配,才能触发取食活动,而对于广食性昆虫来说,许多不同的受体活动特征都能激发取食反应。为了了解取食行为,有必要(a)研究哪些植物化感物质引起接受或拒绝反应,(b)确定参与介导接受或拒绝植物的栓锥感器内的味觉受体细胞的功能和数量,以及(c)描述在这些感器内的受体细胞如何编码这种味觉信息并将其传递给中枢神经系统以阻止或引起取食行为。
6. 植物化学物质和寄主植物偏好
植物化学物质包括初级和次级植物代谢物。次生代谢物质(即植物化感物质)并非普遍存在于高等植物中,而是仅限于某些植物类群(或在这些类群中出现的浓度比其他类群高得多),对昆虫没有营养意义。植物产生广泛的次级代谢物,作为防御食草动物以及微生物。此外,它们还可以作为传粉者的吸引物。还有一些与神经递质有结构上的相似性。许多次级代谢物可能具有细胞毒性,因为它们干扰生物膜、细胞骨架蛋白或DNA,并能诱导细胞凋亡 . 食物的特异性可以完全基于次级代谢物的存在与否。在某些植物类群中,这些化合物也可以作为一些专门的昆虫的 “标志性信号 “刺激,使它们能够明确地识别它们的宿主植物,以及作为对非适应性物种的有效防御屏障。 取食抑制剂(抑制取食的次生植物物质)在宿主植物的相互作用中起着重要作用。有人推测,寄主植物的选择或宿主植物的可接受性是由于缺乏抑制取食的化合物,而对非宿主植物的排斥是由于存在取食抑制剂或阻遏剂。缺乏抑制取食的化合物和拒绝非寄主植物是由于存在取食抑制剂或威慑剂。术语 “植物化感物质”(allelochemic)被创造并定义为由一个物种(植物)的个体产生的,并影响另一个物种(昆虫)的生长、健康、行为或种群的 “非营养化学物”。常见的是,一种植物可能产生不止一种植物化感物质,它们被储存在植物的重要部位。
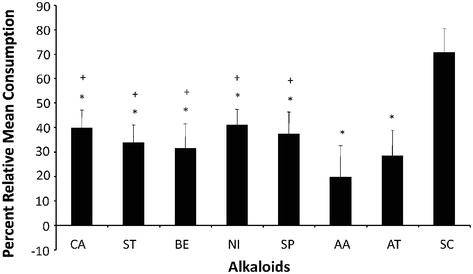
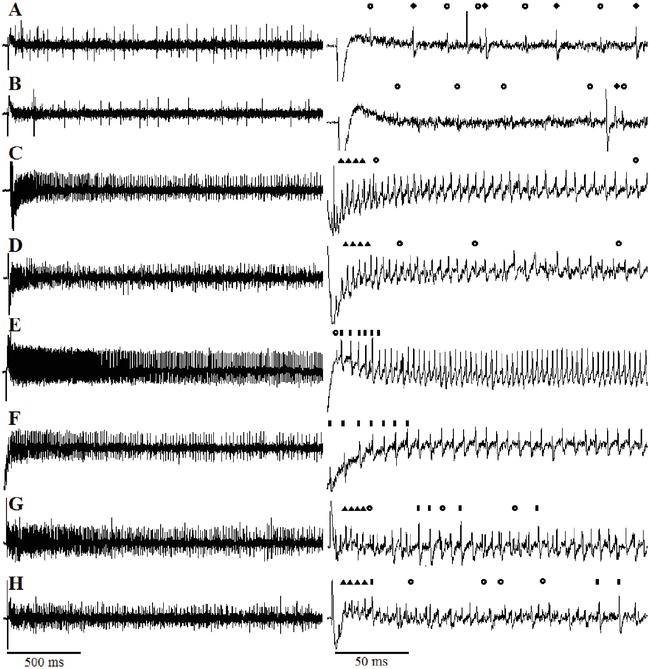
舞毒蛾的幼虫显示出广泛的宿主植物偏好。它们是高度多食性的取食者(以属于不同植物科的许多植物为食)。对于多食性(“多食性”)昆虫物种,如舞毒蛾,可能存在取食刺激物和取食抑制物之间的平衡,这决定了植物将被吃或被拒绝的程度。虽然取食刺激剂是驱动取食的必要条件,但它不可能影响宿主植物的选择。因此,宿主植物的选择可能是由非宿主的抑制性化合物的存在决定的。多食性昆虫被阻止取食储存有毒代谢物的植物,它们通常选择那些毒性代谢物活性较低的植物。它们也可能通过迅速改变寄主植物来避免中毒,并且已经进化出对某些植物化感物质的解毒和快速排泄机制。相反,对于许多寡食性(以几种植物物种为食,属于同一植物科)和所有单食性(只以一种或几种密切相关的植物物种为食)(“专食性昆虫”)昆虫物种,其取食似乎是由作为 “标志性刺激化合物"的化学物质的存在所驱动。这使昆虫能够明确地识别它们的宿主植物并刺激进食,以及作为对非适应性物种的有效防御屏障和识别非宿主植物中存在的抑制性化合物。这些 “标志性刺激化合物"可能最初是有毒的,但可以被容忍(解毒)或被封存,用于昆虫对捕食者的防御,或在宿主植物中显示出相对缺乏的抑制作用。舞毒蛾幼虫是 “通吃 “的,在爆发的年份能够摧毁整片森林。相对来说,很少有研究记录哪些植物化感物质与引起这种多食性昆虫的接受或拒绝进食反应有关。迄今为止,只有一项研究描述了外颚叶栓锥感器内的详细超微结构形态和感觉生理学,该感器被认为是参与取食的主要器官。因此,我们对舞毒蛾幼虫化学感器的基本机制的认识远落后于其他鳞翅目幼虫,如烟草天蛾、大菜粉蝶和家蚕。舞毒蛾幼虫的双向取食选择行为生物测定显示,含有生物碱的植物,是最大的植物化感物质成分之一,出现在20-30%的高等植物中,是舞毒蛾幼虫不喜欢的(图3和4)。

7. 味觉受体细胞分类与外周味觉编码
昆虫和其他动物一样,可以尝到对其发育、生存和繁殖至关重要的主要营养物质,包括糖和无机盐。鳞翅目动物通常使用对各种化学物质敏感的独立的细胞来传递有关化学物质存在的信息,包括糖(“糖敏感细胞”)、肌醇(“肌醇敏感细胞”)、盐(“盐敏感细胞”)和抑制剂(“苦味敏感细胞”)。在人类中,后者的化合物会让人尝到 “苦味”。这种分类不一定意味着这些细胞只对这几组化学化合物有反应,但对它们更敏感,并可能被它们激活。
来自食物的感觉输入引起了昆虫的行为反应。然而,没有直接的实验证据表明,来自味觉受体的输入是如何在中枢神经系统中整合的。然而,取食的数量和味觉感器细胞对不同浓度的刺激物的活动之间似乎有直接关系。相反,当抑制细胞的活性随着抑制物浓度的增加而增加时,取食的量就会下降。据推测,这些输入在中枢神经系统中以相加的方式汇集在一起并产生积极的影响。另一方面,抑制剂对取食有消极影响。昆虫的味觉受体将复杂的植物化学成分的质量和数量转变成动作电位的神经编码。由植物汁液等产生的复杂刺激常常在支配一个或多个感器内的几个受体细胞中引起spike串联。通常情况下,每个细胞类型(例如,糖敏感细胞与苦味敏感细胞)可以根据其spike的振幅和频率加以区分。spike序列中动作电位的频率和时间分布包含了有关刺激的信息。轴突投射到第一中继站–SOG,并在此汇聚,没有中间性的突触。通过分析 “输入-输出 “关系来解开感觉密码。这可以通过刺激特定的感器和量化动作电位的电生理记录(输入),以及根据取食多少食物来量化行为(输出)来实现。编码是通过在输入和输出之间建立关联来推断的。
为了更好地理解化学感受器官和中枢神经系统之间的神经通讯导致的接受或拒绝行为,存在三种理论来描述感觉反应。(1)线性标记模式,(2)纤维交叉模式,和(3)时间模式。第一个理论提出单一化合物在控制或改变行为方面越重要,其感受就越可能由单一细胞编码。这条通往中枢神经系统的 “标记线”(即信息沿线传递到大脑的线或轴突)将只携带来自对特定化合物(或化学物家族)具有狭窄和明确的反应谱的细胞的信息,并将与特定的行为反应直接联系起来。第二种理论认为,神经系统通过评估来自许多具有不同但重叠的反应谱的单个感觉细胞的反应来决定行为输出。中枢神经系统通过阅读和处理所有传入感觉纤维(轴突)的同时输入来提取有意义的信息(纤维交叉模式)。这在脊椎动物中也是已知的。第三种理论,时间模式化可能叠加在跨纤维模式化上,表明不同细胞之间的反应比例随时间变化,并能修改特定的信息。最重要的是,应该注意到这三种理论并不相互排斥,可以结合成一个模型。
介导接受的感觉编码:(i) 刺激编码为接受特征的特定糖细胞;(ii) 刺激中枢神经系统识别为接受特征的广谱糖细胞和(iii) 抑制特定的取食抑制性受体;这有助于接受的神经编码。取食刺激物可通过以下方式改变感觉输入。(i) 刺激特定的抑制性受体;(ii) 刺激广谱受体;(iii) 刺激一些细胞,抑制其他细胞,从而改变复杂而微妙的编码;(iv) 抑制特定的取食刺激性受体;这有助于抑制性化合物的神经编码,以及(v) 引起高度不自然的脉冲模式,往往频率很高。抑制性神经元对多种化学物质的反应能力是由于它有多种多样的受体位点,每个受体位点都有自己的结构-功能特异性,或者是由于活性化学物质具有共同的特征,使得它们能够与单一受体位点相互作用。抑制性细胞具有一些独特的特征。(i)它们通常比对取食刺激化合物有反应的细胞适应得更慢;(ii)取食抑制性受体的强活动比其他类型的细胞稳定在一个较高的水平;(iii)在强反应之前可能有一个相对较长的潜伏期;(iv)在应用刺激后,尖峰频率可能缓慢增加,以及(v)尖峰幅度可能随刺激浓度增加。因此,不同的适应率有助于解释感受编码如何随时间变化,以及当感觉信息被送到大脑时,抑制性受体活动如何逐渐变得更加明显。在一餐开始时可能是可以接受的食物,很快就会变得不可接受,因为在总的感觉印象中,抑制力的份额更加突出。以大菜粉蝶为模型,确定来自传递抑制信息的受体细胞的冲动被中枢神经系统赋予更大的权重。因此,来自抑制敏感的神经元的一个冲动可能会中和来自糖敏感细胞的2.5个冲动。此外,信号显示有植物化感物质存在的细胞通常对比测量营养物质数量的受体低约1000倍的浓度作出反应。

8. 结论
昆虫是研究支配取食行为的机制的理想模型。如前所述,鳞翅目幼虫的味觉感器为解决有关味觉系统的问题提供了一个很好的系统。这些感器形成了一个i)相对简单的感觉系统,有少量的感觉细胞介导味觉机制,ii)这些感器的位置为实验操作提供了相对容易的途径,以及iii)这些感器内的受体细胞是可以单独识别的,并表现出典型的可重复的电生理反应。在脊椎动物和无脊椎动物(即昆虫)之间,味觉的解剖组织和分子信号传导途径明显不同。然而,在这两个动物群体中,味觉质量的编码显示出惊人的相似性,如每一种味觉质量都是由一个标记线所介导的。这意味着一个特定的味觉感受细胞群被分离出来,负责编码特定的味觉质量。
在分子水平上,最近对家蚕的研究表明,三种昆虫味觉受体(GRs)(BmGr16、BmGr18和BmGr53)对结构不同且部分重叠的抑制性化合物反应广泛,表明这些苦味GRs是取食抑制性化合物受体,在宿主植物识别中发挥重要作用。有趣的是,对家蚕的取食偏好研究表明,GRr66,是这种单食性昆虫对桑树的取食偏好的原因。借助于CRISPR-Cas9系统,在Gr66基因座上引入了一个突变。作为这一基因突变的结果,家蚕幼虫扩大了它们的取食范围。幼虫以几种通常不在其食物中的植物物种为食,从而发现了第一个遗传和表型证据,证明单一的苦味GR可以影响这种昆虫的取食偏好。最近在功能基因组学方面的进展和关于家蚕的苦味GRs的分子进展,指出了控制虫害的新方向和策略。此外,它扩大了我们对昆虫与植物相互作用的理解,并产生了关于昆虫如何感知和处理味觉信息的新思路。
参考
DOI: http://dx.doi.org/10.5772/intechopen.99293
License
Copyright 2021 Guo-Cheng Li. Released under the MIT license.
Guo-Cheng Li
PhD of cell biology
Hi! I am a PhD at Institute of Zoology, The Chinese Academy of Sciences, Beijing, China. I am using genomics and transcriptomics to study insect chemoreception.